Download præsentationen
Præsentation er lastning. Vent venligst

1
T celle medierede funktioner
T LYMFOCYTTER udvikling og modning T celle medierede funktioner Marianne Hokland

2
naïve T lymfcocytter forlader thymus -> cirkulationen

3
RESULTATET - so far udvikling af naïve T lymfocytter, der.. ---
- udtrykker TCRgamma/delta (ca 5%) eller - TCRalpha/beta (95%) der udtrykker enten CD4 (2/3) eller CD8 (1/3) og som genkender fremmede antigener præsenterede på egne MHC molekyler og som IKKE reagerer på egne peptider
 eller. - TCRalpha/beta (95%) der udtrykker enten CD4 (2/3) eller CD8 (1/3) og som genkender fremmede antigener præsenterede på egne MHC molekyler. og som IKKE reagerer på egne peptider.")
4
cirkulerende naïve t celler
TCRαβ (>95%) CD4+ (Thelper) - ca 2/3 CD8+ (Tcytotox) - ca 1/3 Monocytes are WBCs that are typically round with a horseshoe-shaped nucleus. They are released into the blood and then migrate into tissues where they become macrophages. Functionally, they are phagocytic. Some macrophages simply ingest and destroy foreign material, others ingest, digest and then display fragments of the foreign material on MHC molecules (see notes for slide #8, “T cell training”. These macrophages are then called antigen presenting cells or APCs. TCRγδ (<5%) 20-30% lymfocytter i PB heraf er godt 80% T lymfocytter
 CD4+ (Thelper) - ca 2/3. CD8+ (Tcytotox) - ca 1/3. Monocytes are WBCs that are typically round with a horseshoe-shaped nucleus. They are released into the blood and then migrate into tissues where they become macrophages. Functionally, they are phagocytic. Some macrophages simply ingest and destroy foreign material, others ingest, digest and then display fragments of the foreign material on MHC molecules (see notes for slide #8, T cell training . These macrophages are then called antigen presenting cells or APCs. TCRγδ (<5%) 20-30% lymfocytter i PB. heraf er godt 80% T lymfocytter.")
5
T celle funktioner - hovedtræk
CD4 CD8 ELIMINATION AF INTRACELLULÆRE MIKROORGANISMER

6
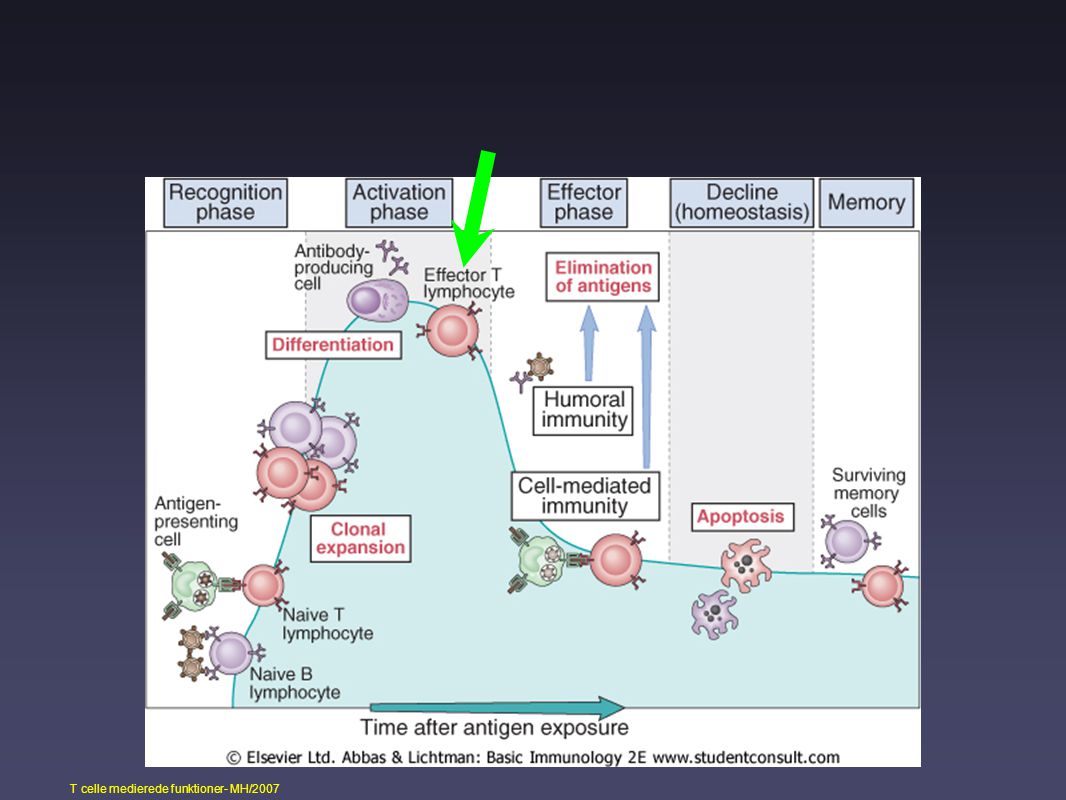
“Life story of the T cell”
cirkulering (“patruljering”) primær aktivering (i lymfeknude) effektorfunktion(er) “dæmpning” af response/homeostase memory
 primær aktivering (i lymfeknude) effektorfunktion(er) dæmpning af response/homeostase. memory.")
7
adaptivt immunrespons oversigt

8
AGENDA cirkulering (“patruljering”) primær aktivering (i lymfeknude) effektorfunktion(er) “dæmpning” af response/homeostase memory

9
T lymfocyt traffik hvordan sikres, at T lymfocytterne kan checke - og reagere på - infektion overalt i kroppen?

10
T lymfocyt traffik AV

11
lymfocyt traffik Ag “naive” T lymfocytter ductus thoracicus
fra ductus thoracicus -> vena cava inferior - gennem hjertet og på den igen ductus thoracicus

12
TRANSPORT AF ANTGEN TIL LYMFEKNUDER

13
ANTIGEN “OPSAMLING” fig 3.2

14
T lymfocytter migrerer ind i lymfeknuden

15
HEV - T LYMFOCYTTENS “INDGANGSPORT”

16
T lymfocytter migrerer ind i lymfeknuden
venoler - karret udvider sig - og blodstrømmen bliver derfor langsommere - giver bedre muligheder for at lymfocytterne kan “rulle” - og senere trænge ind

17
T LYMFOCYT MIGRATION GENNEM HEV

18
T celle migration ind i LN
det tager lymfocytten 6-12 timer at migrere igennem en lymfeknude - dvs den kan cirkulere et par gange i døgnet de T celler, der ikke “møder” deres antigen i LN migrerer ud igen

19
MIGRATION I LN når de naïve T celler er trængt ind igennem HEV “skanner” de lymfeknuden for antigener (på APC) - det tager ca 12 timer dvs - de naïve T celler kan cirkulere ca 2 gange i døgnet

20
AGENDA cirkulering (“patruljering”) primær aktivering (i lymfeknude)
effektorfunktion(er) “dæmpning” af response/homeostase memory
 dæmpning af response/homeostase. memory.")
21
FØRSTE led i T celle aktiveringen er
Antigen præsentation FØRSTE led i T celle aktiveringen er ANTIGEN PRÆSENTATION

22
lymfeknuden “opsamler” antigener fra mikroorganismer
- og den foregår i sekundære lymfoide organer - specielt LYMFEKNUDER og MILT

23
ANTIGEN PRÆSENTATION (genkendelse)
")
24
T celle migration ind i LN
de T celler, der “møder” deres antigen i LN gennemgår proliferation og aktivering

25
Aktivering af naïve T celler
Fig 5.3 (udsnit)
")
26
aktivering af naïve T celler
Den første betingelse som SKAL opfyldes før T cellen ændrer adfærd er binding imellem TCR og et MHC molekyle hvortil der er bundet et peptid. Peptidet stammer fra en processering af materiale som APC’en har opsamlet i de perifere væv. De fleste af de T celler som genkender peptider fra vore egne proteiner er sorteret fra i Thymus under den negative selektion. Derfor er en binding imellem et MHC-peptid complex og en TCR et tegn på at der er fremmed materiale tilstede i kroppen. Bindingen er dog ikke et entydigt tegn. Der kan være tale om MHC-TCR binding imellem T celler som har undveget den negative selektion. Men frygt ej. Hertil har immun forsvaret et sikkerhedsnet. Udover TCR binding skal T cellen også modtage et co-stimulatorisk signal. Dette leveres af APC’en, men kun hvis APC’en har registreret tilstedeværelsen af fremmed materiale. Hvilken adfærd denne kontakt medfører afhænger af sekundære og tertiære signaler som T cellen primært modtager fra APC’en. Disse signaler kan være i form af co-stimulatoriske membran-bundne og/eller secernerede ligander. fig 5.4

27
aktivering af naïve T celler

28
DEN IMMUNOLOGISKE SYNAPSE

29
T cellen behøver mere end ét signal
- det 3. signal “bestemmer” i princippet, hvilken T lymfocyt subpopulation der udvikles

30
hvis IKKE mindst 2 signaler ingen effekt eller anergi
hvorfor er det smart?? - ja - hvad er det nu der bestemmer, om den antigenpræsenterende celle udtrykker co-receptorer?? TLR aktivering af mikroorganismer på DC - det er bla noget af det, cancerpatienter mangler!!

31
binding “omdannes” til et biokemisk signal
Biokemiske enzymer (typisk kinaser) og substrater er associeret med cyotoplasmiske domæner på forskellige overflade associerede proteiner Ved dannelse af den immunologiske synapse bringes enzymer og substrater sammen Enzymer har nu i meget højere grad adgang til deres substrater end hvis den immunologiske synapse ikke er dannet Resultatet er at phosphorylerede substrater dannes i meget højere grad end normalt, hvilket kan initiere en signalerings kaskade Hos T celler vil signalerings kaskaden som initieres af dannelsen af den immunologiske synapse kunne resultere i cytokin produktion og/eller proliferation
 og substrater er associeret med cyotoplasmiske domæner på forskellige overflade associerede proteiner. Ved dannelse af den immunologiske synapse bringes enzymer og substrater sammen. Enzymer har nu i meget højere grad adgang til deres substrater end hvis den immunologiske synapse ikke er dannet. Resultatet er at phosphorylerede substrater dannes i meget højere grad end normalt, hvilket kan initiere en signalerings kaskade. Hos T celler vil signalerings kaskaden som initieres af dannelsen af den immunologiske synapse kunne resultere i cytokin produktion og/eller proliferation.")
32
aktivering

33
Klonal proliferation Aktivering af T cellen følges umiddelbart af klonal expansion. T cellerne deler sig hurtigt og opbygger en “hær” af celler som kan bekæmpe den invaderende microbe Klonerne har meget kort levetid...omkring et par dage. Ud over binding til TCR skal T cellen modtage mindst ét co-stimulatorisk signal for at blive aktiveret. CD28 er et klassisk eksempel på en costimulatorisk receptor. Mindre kendte er ICOS og SLAM. CD28 bindes til overflade protein B7 på APC’en B7 opreguleres ved aktivering T celler som aktiveres uden at modtage co-stimulatorisk signal bliver inaktive (anergiske) (CD4 og CD8 er IKKE costimulatoriske signaler. De er co-receptorer for TCR complexet) hvorfor er klonal proliferation vigtig???? det er dynamikken i immunsystemet - eller om I vil - den måde immunsystemet adapterer til den aktuelle situation (eks infektion) Antallet af antigen specifikke celler kan stige fra ca. 1 ud af en million inden infetion til omkring 1 ud af 50 efter klonal expansion.
 (CD4 og CD8 er IKKE costimulatoriske signaler. De er co-receptorer for TCR complexet) hvorfor er klonal proliferation vigtig det er dynamikken i immunsystemet - eller om I vil - den måde immunsystemet adapterer til den aktuelle situation (eks infektion) Antallet af antigen specifikke celler kan stige fra ca. 1 ud af en million inden infetion til omkring 1 ud af 50 efter klonal expansion.")
34
T lymfocyt aktivering Fig 5.2

36
T lymfocyt subpopulationer

37
B celle hjælp CD4+ T lymfocytter hjælper T celler i forb med differentiering og antistofproduktion - - bestemmer isotypeswitch - og somatisk mutation

38
B celle hjælp

39
B celle hjælp

40
T “hjælper” celle populationer

41
Th1 lymfocytter

42
Th2 lymfocytter

43
Th17 lymfocytter

44
Treg lymfocytter

45
Cytotoxisk T lymfocyt drab

46
Cytotoxisk T lymfocyt drab

47
Cytotoxisk T lymfocyt drab

48
Elimination af intracellulære mikroorganismer

49
T lymfocyt homeostase

50
effektorfase!!

51
MISSION ACCOMPLISHED!!

Lignende præsentationer






>")












